Вызванная активность мозга при восприятии фигур Канизса
Аннотация
В статье рассматриваются проблемы зрительного восприятия целостного объекта. В качестве объективного метода, позволяющего исследовать данные вопросы, используются в качестве стимулов фигуры, формирующих субъективные контуры - фигуры Канизса. В обследовании приняли участие 8 практически здоровых мужчин (студенты и сотрудники ЮФУ), правшей, в возрасте от 22 до 35 лет. Результаты дисперсионного анализа характеристик зрительных ВП, зарегистрированных при восприятии различных фигур, показали, что значимые различия обнаруживались при сравнении реальной и иллюзорной форм с фигурой контроля.
Ключевые слова: Зрительное восприятие, фигуры Канизса, вызванные потенциалы (ВП)
05.11.17 - Приборы, системы и изделия медицинского назначения
Изучение механизмов зрительного восприятия целостного объекта остается одной из наиболее актуальных проблем как нейро, так и психофизиологии. Одним из возможных механизмов связывания отдельных элементов воспринимаемого образа в целостную картину, т.е., «перцептивного связывания» (binding problem), является, как полагают, синхронизация активности нейронных ансамблей мозга в различных частотных диапазонах. Эффективным методическим приемом, позволяющим исследовать вопросы, связанные с восприятием целостного объекта, является использование в качестве стимулов фигур, формирующих субъективные контуры (фигуры Канизса). Как было показано в работах отечественных и зарубежных авторов, восприятие таких фигур сопровождается изменением активности в различных областях мозга [1- 5].
Задачей нашего исследования было исследование взаимосвязи процессов восприятия фигур Канизса с параметрами зрительных ВП. В обследовании приняли участие 8 практически здоровых мужчин (студенты и сотрудники ЮФУ), правшей, в возрасте от 22 до 35 лет. Все они имели нормальное или скорректированное до нормального зрение. В качестве зрительных стимулов использовались модифицированный треугольник Канизса, реальный треугольник и не треугольник. Стимулы предъявлялись на экране монитора, расположенного на расстоянии около 1 м на уровне глаз обследуемого. Время экспозиции каждого стимула – 200 мс, межстимульный интервал – от 2 до 5 с, размер фигур - 7 угловых градусов. Момент предъявления был синхронизован с кадровой разверткой монитора. Обследуемый располагался в кресле, в удобной позе, в звуко – изолированной камере. Ему предлагалось фиксировать взгляд в центре экрана, где находилась точка фиксации (крест). Длительность обследования составляла 20 минут.
ЭЭГ регистрировали с помощью хлорсеребряных электродов непрерывно от 6 отведений, расположенных по системе 10-20% (P3, P4, Т5, Т6, O1, O2), с использованием усилителя биопотенциалов Нейровизор БММ («NEUROBOTICS», Москва). В качестве референтного использовался объединенный ушной электрод. Частота квантования сигналов по каждому из каналов составляла 1000 Гц. Частота пропускания усилителей составляла 0,1-300 Гц. Контроль глазных движений осуществляли с помощью регистрации ЭОГ.
После фильтрации ЭЭГ (удалялись частоты до 1 Гц и 50 Гц), для анализа ВП отбирались безартефактные отрезки электрограмм, в которых отфильтровывались частоты свыше 20 Гц. Эпоха анализа составляла 1024 мс, предстимульный интервал – 128 мс. ВП получали путем усреднения эпох для каждого типа стимулов – иллюзорный треугольник, реальный треугольник, не треугольник.
Путем усреднения ответов, зарегистрированных у всех обследуемых, для каждого типа стимула получались обобщенные ВП (Grand Average), на которых были идентифицированы следующие компоненты: P100 (ЛП70 - 120мс), N150 (125-175 мс), P200 (190-235мс), N400 (350 - 450мс). Индивидуальные ВП рассчитывались для каждого обследуемого отдельно.
Статистическую оценку амплитуд (А) и латентных периодов (ЛП) компонентов ВП осуществляли с использованием многофакторного дисперсионного анализа (ANOVA) из пакета прикладных программ Statistica 5. При сравнении выделяли следующие факторы: тип стимула (Т, уровни: иллюзорный треугольник, реальный треугольник, не треугольник), отведения (L, уровни: P3, P4, Т5, Т6, O1, O2), компоненты (Р, уровни: P100, N150, P200, N400). При величине p ≤ 0,05 различия рассматривались как статистически достоверные, при 0,05 ≤ p ≤ 0,08 – как существенные (констатировалось наличие жесткого тренда).
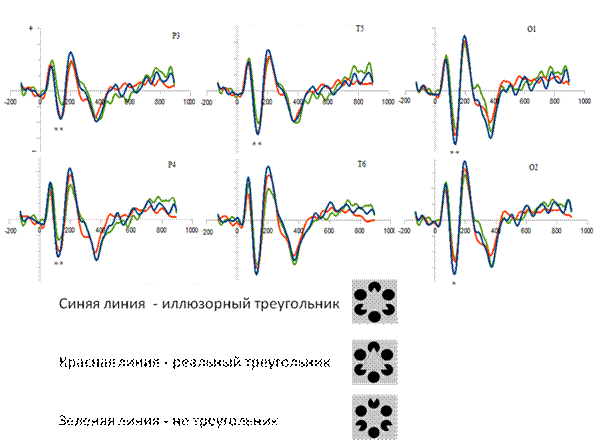
Рис. 1. Графическое изображение усредненных ЗВП (Grand average), зарегистрированных в ЭЭГ затылочных (О1, О2), теменных (Р3, Р4), височных (Т5, Т6) областей коры в ответ на предъявление различных изображений. По оси ординат – амплитуда ЗВП, мкВ; по оси абсцисс – время, мс; **- статистически значимые различия при р ≤ 0,05, * - наличие тренда 0,05 ≤ p ≤0,08.
Результаты дисперсионного анализа характеристик зрительных ВП, зарегистрированных при восприятии различных фигур, показали, что значимые различия обнаруживались при сравнении реальной и иллюзорной форм треугольника с фигурой контроля (не треугольник). При восприятии треугольников (реального и иллюзорного), по сравнению с контролем, А компонента N150 была ниже практически во всех отведениях, а статистически значимо – в затылочных (FO1(1,5)= 8,99 p=0,03; FO2(1,5)=5,99 p=0,058). При сравнении реального треугольника и не треугольника различия обнаруживались в активности симметричных затылочных (FO1(1,5)=6,71 p=0,048; FO2(1,5)= 5,66 p=0,063), левого височного (FT5(1,5)=6,99 p=0,045) и симметричных теменных (FP3(1,5)=10,74 p=0,022; FP4(1,5)=11,35 p=0,02) отведений (Рис.1).
Полученные нами результаты, в целом, совпадают с данными других авторов [1, 2, 5], исследовавших зрительные ВП при восприятии фигур с субъективными контурами и показавших чувствительность компонента N150 к наличию или отсутствию контуров. Различия в А компонентов ВП, регистрируемых в ответ на предъявление стимулов, имеющих и не имеющих контуры, обнаружены нами в активности симметричных теменных, затылочных и левой височной области. В отличие от работы Коршуновой Т.Г. (1998), в которой высказано предположение о наличии в системе зрительного восприятия двух областей (затылочная и левая теменная), кодирующих иллюзорные контуры и фигуры, нами получены данные, указывающие на участие в этом процессе и теменных областей. Полученные нами результаты коррелируют со сведениями, полученными D.Stanley (2003) с помощью функциональной МРТ и указывающими на то, что восприятие субъективных контуров сопровождается активацией латерального затылочного комплекса (ЛОК). Тем не менее, учитывая полученные нами результаты, представляется необходимым дальнейшие исследования мозговой активности, вызванной восприятием фигур Канизса, не только в низкочастотной части спектра ЭЭГ, но и в диапазоне гамма-частот.
Литература:
1.Коршунова С.Г. Зрительные вызванные потенциалы на иллюзорные контуры (Квадрат Канизса) //Журн. высш. нервн. деятельности. - 1998.- Т.48. - N.5. - С. 807- 814.
2.Brodeur M., Lepore F., Lepage M., Bacon B.A., Jemel B., Debruille J.B. Alternative mode of presentation of Kanizsa figures sheds new light on the chronometry of the mechanisms underlying the perception of illusory figures //Neuropsychologia.- 2008. - Vol.46. - P.554–566.
3.Stanley D.A. and Rubin N. fMRI activation in response to illusory contours and salient regions in the human lateral occipital complex//Neuron. - 2003. - Vol.37.- P.323–331.
4.Tallon-Baudry C. Bertrand O., Delpuech C., Pernier J. Stimulus specificity of phase-locked and non-phase-locked 40Hz visual responses in human// J. Neuroscience. - 1996. - Vol.16. - P.4240-4249.
5.Wu X., Zhang D. Early induced beta/gamma activity during illusory contour perception// Neuroscience Letters.- 2009.- Vol.462.- P. 244-247.